|

by
Chris P. McKay
NASA Ames Research Center
September 14, 2004
DOI: 10.1371/journal.pbio.0020302
from
PlosBiology Website
I need a “tricorder”—the convenient, hand-held device featured on
Star Trek that can detect life forms even from orbit. Unfortunately,
we don't have a clue how a tricorder might work, since life forms
don't seem to have any observable property that distinguishes them
from inanimate matter. Furthermore, we lack a definition of life
that can guide a search for life outside Earth. How can we find what
we can't define?
An answer may lie in the observation that life uses
a small, discrete set of organic molecules as basic building blocks.
On the
surface of Europa and in the
subsurface of
Mars, we can
search for alien but analogous patterns in the organics.
Life As We
Know It
The obvious diversity of life on Earth overlies a fundamental
biochemical and genetic similarity. The three main polymers of
biology—the nucleic acids, the proteins, and the polysaccarides—are
built from 20 amino acids, five nucleotide bases, and a few sugars,
respectively.
Together with lipids and fatty acids, these are the
main constituents of biomass: the hardware of life (Lehninger 1975,
p 21). The DNA and RNA software of life is also common, indicating
shared descent (Woese 1987). But with only one example of life—life
on Earth—it is not all that surprising that we do not have a
fundamental understanding of what life is. We don't know which
features of Earth life are essential and which are just accidents of
history.
Our lack of data is reflected in our attempts to define life.
Koshland (2002) lists seven features of life:
(1) program (DNA)
(2)
improvisation (response to environment)
(3) compartmentalization
(4) energy
(5) regeneration
(6) adaptability
(7) seclusion
(chemical control and selectivity)
A simpler definition is that
life is a material system that undergoes reproduction, mutation, and
natural selection (McKay 1991). Cleland and Chyba (2002) have
suggested that life might be like water, hard to define phenomenologically, but easy to define at the fundamental level. But
life is like fire, not water—it is a process, not a pure substance.
Such definitions are grist for philosophical discussion, but they
neither inform biological research nor provide a basis for the
search for life on other worlds.
The simplest, but not the only, proof of life is to find something
that is alive. There are only two properties that can determine if
an object is alive: metabolism and motion. (Metabolism is used here
to include an organism's life functions, biomass increase, and
reproduction.) All living things require some level of metabolism to
remain viable against entropy. Movement (either microscopic or
macroscopic) in response to stimuli or in the presence of food can
be a convincing indicator of a living thing. But both metabolism
(fire) and motion (wind) occur in nature in the absence of biology.
The practical approach to the search for life is to determine what
life needs. The simplest list is probably: energy, carbon, liquid
water, and a few other elements such as nitrogen, sulfur, and
phosphorus (McKay 1991). Life requires energy to maintain itself
against
entropy, as does any self-organizing open system. In the
memorable words of Erwin Schrödinger (1945), “It feeds on
negative
entropy.” On Earth, the vast majority of life forms ultimately
derive their energy from sunlight.
The only other source of primary
productivity known is chemical energy, and there are only two
ecosystems known, both methanogen-based (Stevens and McKinley 1995; Chapelle et al. 2002), that rely exclusively on chemical energy
(that is, they do not use sunlight or its product, oxygen).
Photosynthetic organisms can use sunlight at levels below the level
of sunlight at the orbit of Pluto (Ravens et al. 2000); therefore,
energy is not the limitation for life.
Carbon, nitrogen, sulfur, and
phosphorus are the elements of life, and they are abundant in the
Solar System. Indeed, the Sun and the outer Solar System have more
than 10,000 times the carbon content of the bulk of Earth (McKay
1991). When we scan the other worlds of our Solar System, the
missing ecological ingredient for life is liquid water. It makes
sense, then, that the search for liquid water is currently the first
step in the search for life on other worlds. The presence of liquid
water is a powerful indication that the ecological prerequisites for
life are satisfied.
Orbital images, such as the canyon in Figure 1 below, show clear evidence
of the stable and repeated, if not persistent, flow of a
low-viscosity fluid on Mars at certain times in its past history.
The fluid was probably water, but the images could also suggest
wind, ice, lava, even carbon dioxide or sulfur dioxide. Recently,
results from the Mars Exploration Rover missions have shown that
this liquid carried salts and precipitated hematite in concretions.
The case for water, we could say, is tight.
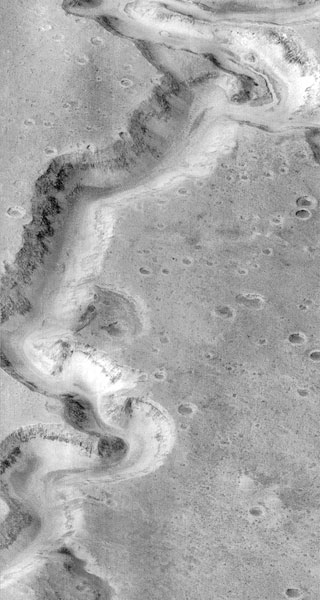
Figure 1. Water on
Another World
.
A Mars Global
Surveyor image showing Nanedi Vallis in the Xanthe Terra region of
Mars.
The image covers an
area 9.8 km ×18.5 km; the canyon is about 2.5 km wide.
This image is the
best evidence we have of liquid water anywhere outside the Earth.
Photo credit: NASA/Malin
Space Sciences.
On Jupiter's moon Europa, the cracks and
icebergs on the surface of the ice indicate water beneath the ice,
but not necessarily at the present time. Present water on Europa is
indicated by the magnetic disturbance Europa makes as it moves
through Jupiter's magnetic field, not unlike the way coins in the
pocket of a passenger will set off an airport metal detector. Europa
has a large conductor, and this is most likely a global, salty layer
of water.
Viking on
Mars: Been There, Tried That
The Viking missions to Mars in the late 1970s were the first (and as
yet, the only) search for life outside Earth. Each Viking conducted
three incubation experiments to detect the presence of metabolism in
the Martian soil. Each lander also carried a sophisticated Gas
Chromatograph Mass Spectrometer for characterizing organic
molecules. The results were unexpected (Klein 1978, 1999). There was
a detectable reaction in two of the incubation experiments. In the
“Gas Exchange” experiment, a burst of oxygen was released when the
soil was exposed to water.
The “Labeled Release” experiment showed
that organic material was consumed, and that carbon dioxide was
released concomitantly. In the Labeled Release experiment, this
reaction ceased if the soil was first heated to sterilizing
temperatures, but the reaction of the Gas Exchange Experiment
persisted.
If considered alone, the Labeled Release results would be a
plausible indication for life on Mars. However, the Gas
Chromatograph Mass Spectrometer did not detect the presence of any
organic molecules in the soil at level of one part per billion (Biemann
1979). It is difficult to imagine life without associated organic
material, and this is the main argument against a biological
interpretation of the Viking results (Klein 1999; but cf. Levin and
Straat 1981).
It is also unlikely that the oxygen release in the
Gas
Exchange experiment had a biological explanation, because the
reaction was so rapid and persisted after heating. It is generally
thought that the reactivity observed by the Viking biology
experiments was caused by one or more inorganic oxidants present in
the soil, and was ultimately produced by ultraviolet light in the
atmosphere. Consistent with the apparently negative results of the
Viking biology experiments, the surface of Mars also appears to be
too dry for life. Indeed, conditions are such that liquid water is
rare and transient, if it occurs at all (e.g., Hecht 2002).
It's Life,
Jim, but Not As We Know It
Table 1 below, shows a categorization of life as we have observed it. Using
this diagram, we can speculate about how life might be different on
Mars or Europa. At the bottom of the table, life is composed of
matter—a reasonable assumption for now. Carbon and liquid water are
the next level; this makes Mars and Europa likely candidates,
because they have carbon and have, or have had, liquid water. Other
worlds may have a different chemical baseline for life.
The usual
speculation in this area is that the presence of ammonia and
silicon, rather than water and carbon, might be preconditions for
life on other planets. Such speculation has yet to lead to any
specific suggestions for experiments, or to new ways to search for
such life, but this may just reflect a failure of human imagination
rather than a fundamental limitation on the nature of life.
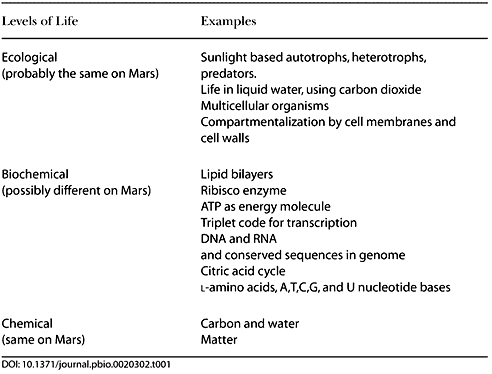
Table 1. A
Categorization of Structures That Comprise Terrestrial Life
Life on Mars is also likely to be the
same at the top of the table: at the ecological level. Primary
production in a Martian ecosystem is likely to be phototrophic,
using carbon dioxide and water. Heterotrophs are likely to be
present to consume the phototrophs and in turn to be consumed by
predators. Darwinian evolution would result in many of the same
patterns we see in ecosystems on Earth.
While it may be similar at
the top (ecological) and bottom (chemical) levels, life on Mars
could be quite alien in the middle, in the realm of biochemistry.
Pace (2001) has argued that alien biochemistry will turn out to be
the same as biochemistry on Earth, because there is one best way to
do things and that natural selection will ensure that life
everywhere discovers that way. Only observation will tell if there
is one possible biochemistry, or many.
Future missions to Mars might find microfossils in sedimentary rocks
such as those at Meridiani Planum. Microbes don't readily form
convincing fossils; the one exception may be the strings of
magnetite formed by magnetotactic bacteria (Friedmann et al. 2001).
As interesting as fossils might be, we could not be sure that a
fossil found on Mars was not merely another example of Earth life.
We know that rocks have come to Earth from Mars, and it is possible
that such rocks could have carried life between the planets (Mileikowsky
et al. 2000; Weiss et al. 2000). Finding fossil evidence for life on
Mars does not demonstrate a second genesis in our Solar System.
Finding a Way
to Search for Alien Life
If we were to find organic material in the subsurface of Mars or on
the ice of Europa, how could we determine whether it was the product
of a system of biology or merely abiotic, organic material from
meteorites or photochemistry? If this life were in fact related to
Earth life, this should be easy to determine. We now have very
sensitive methods, such as PCR and fluorescent antibody markers, for
detecting life like us. This case would be the simplest to
determine, but it would also be the least interesting. If the life
turned out to be truly alien, then the probes specific to our
biology would be unlikely to work. What, then, could we do to
determine a biological origin?
The question is open and possibly urgent. On space missions already
being planned, we may have the opportunity to analyze the remains of
alien organics on the surface of Europa or frozen below ground on
Mars. The instruments that will make this analysis must be designed
in the next couple of years.
One approach appears promising. I call it the “Lego Principle.” It
is based on the patterns of the molecules of life. Biological
processes, in contrast to abiotic mechanisms, do not make use of the
range of possible organic molecules. Instead, biology is built from
a selected set. Thus, organic molecules that are chemically very
similar to each other may have widely different concentrations in a
sample of biological organics. An example of this on Earth is the 20
amino acids used in proteins and the use of the left-handed version
of these amino acids.
The selectivity of biological processes is
shown schematically in Figure 2 by the distribution of spikes in
contrast to a smooth, nonbiological distribution. General arguments
of thermodynamic efficiency and specificity of enzymatic reactions
suggest that this selectivity is required for biological function
and is a general result of natural selection. Different life forms
are likely to have different patterns, and at the very least we
might find the mirror symmetry of life on Earth, with d- instead of
l-amino acids.
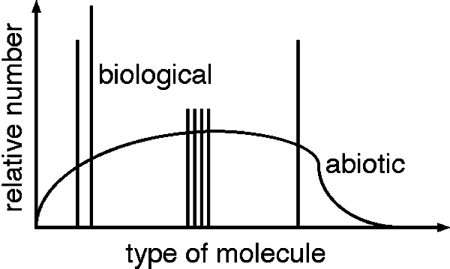
Figure 2. Comparison
of Biogenic with Nonbiogenic Distributions of Organic Material
.
Nonbiological
processes produce smooth distributions of organic material,
illustrated here by the curve.
Biology, in contrast
selects and uses only a few distinct molecules, shown here as spikes
(e.g., the 20 l-amino acids on Earth).
Analysis of a sample
of organic material from Mars or Europa may indicate a biological
origin if it shows such selectivity.
This approach has immediate practical
benefit in the search for biochemistry in the Solar System. Samples
of organic material collected from Mars and Europa can be easily
tested for the prevalence of one chirality of amino acid over the
other. More generally, a complete analysis of the relative
concentration of different types of organic molecules might reveal a
pattern that is biological even if that pattern does not involve any
of the familiar biomolecules.
Interestingly, if a sample of organics
from Mars or Europa shows a preponderance of D-amino acids, this
would be evidence of life, and at the same time would show that this
life was distinct from Earth life. This same conclusion would apply
to any clearly biological pattern that was distinct from that of
Earth life.
Organic material of biological origin will eventually lose its
distinctive pattern when exposed to heat and other types of
radiation, (examples of this include the thermal racemization of
amino acids), but at the low temperatures in the Martian permafrost,
calculations suggest that there has been no thermal alteration (Kanavarioti
and Mancinelli 1990). An interesting question, as yet unanswered, is
how long organic material frozen into the surface ice of Europa
would retain a biological signature in the strong radiation
environment.
On Europa, the organic material for our tests might be collected
right from the dark regions on the surface. On Mars, there is
ice-rich ground in the cratered southern polar regions (Feldmann et
al. 2002), which presumably overlies deeper, older ice. The surprise
discovery of strong magnetic fields in the southern hemisphere of
Mars (Acuña et al. 1999; Connerney et al. 1999) indicates that the
area may be the oldest undisturbed permafrost on that planet.
Like
the mammoths extracted from the ice in Siberia, any Martian microbes
found in this ice would be dead, but their biochemistry would be
preserved. From these biological remains, it would then be possible
to determine the biochemical composition of, and the phylogenetic
relationship between, Earth life and Martian life. We may then have,
for the first time, a second example of life.
References
-
Acuña MH, Connerney JEP, Ness NF,
Lin RP, Mitchell D, et al. (1999)
Global distribution of crustal magnetism
discovered by the Mars Global Surveyor MAG/ER experiment.
Science 284: 790–793.
-
Biemann K (1979)
The implications and limitations of the
findings of the Viking Organic Analysis Experiment. J
Mol Evol 14: 65–70.
-
Chapelle FH, O'Neil K, Bradley PM,
Methé BA, Ciufo SA, et al. (2002)
A hydrogen-based subsurface microbial
community dominated by methanogens. Nature 415:
312–315.
-
Cleland CE, Chyba CF (2002) Defining
life.
Orig Life Evol Biosph 32: 387–393.
-
Connerney JEP, Acuna MH, Wasilewski
P, Ness NF, Reme H, et al. (1999)
Magnetic lineations in the ancient crust
of Mars. Science 284: 794–798.
-
Feldman WC, Boynton WV, Tokar RL,
Prettyman TH, Gasnault O, et al. (2002)
Global distribution of neutrons from Mars:
Results from Mars Odyssey. Science 297: 75–78.
-
Friedmann EI, Wierzchos J, Ascaso C,
Winklhofer M (2001)
Chains of magnetite crystals in the
meteorite ALH84001: Evidence of biological origin.
Proc Natl Acad Sci U S A 98: 2176–2181.
-
Hecht MH (2002)
Metastability of liquid water on Mars.
Icarus 156: 373–386.
-
Kanavarioti A, Mancinelli RL (1990)
Could organic matter have been preserved on Mars for 3.5 billion
years? Icarus 84: 196–202.
-
Klein HP (1978)
The Viking
biological experiments on Mars. Icarus 34: 666–674.
-
Klein HP (1999)
Did Viking discover
life on Mars? Orig Life Evol Biosph 29: 625–631.
-
Koshland DE Jr (2002)
The seven
pillars of life. Science 295: 2215–2216.
-
Lehninger AL (1975) Biochemistry.
>New York: Worth. 1104 p.
-
Levin GV, Straat PA (1981)
A search
for nonbiological explanation of the Viking labeled release life
detection experiment. Icarus 45: 494–516.
-
McKay CP (1991) Urey Prize lecture:
Planetary evolution and the origin of life. Icarus 91: 92–100.
-
McKay CP (1997)
The search for life
on Mars. Orig Life Evol Biosph 27: 263–289.
-
Mileikowsky C, Cucinotta FA, Wilson
JW, Gladman B, Horneck G, et al. (2000)
Natural transfer of
viable microbes in space. Icarus 145: 391–427.
-
Pace N (2001) The universal nature
of biochemistry. Proc Natl Acad Sci U S A 98: 805–808.
-
Raven JA, Kübler JE, Beardall J (2000)
Put out the light, and
then put out the light. J Mar Biol Ass (UK) 80: 1–25.
-
Schrödinger E (1945) What is life?
Cambridge: Cambridge University Press. 91 p.
-
Stevens TO, McKinley JP (1995)
Lithoautotrophic microbial ecosystems in deep basalt aquifers.
Science 270: 450–454.
-
Ultee A, Souvatzi N, Maniadi K,
König H (2004)
Identification of the culturable and
nonculturable bacterial population in ground water of a
municipal water supply in Germany. J Appl Microbiol 96: 560–568.
-
Weiss BP, Kirschvink JL,
Baudenbacher FJ, Vali H, Peters NT, et al. (2000)
A low
temperature transfer of ALH84001 from Mars to Earth. Science
290: 791–795.
-
Woese CR (1987)
Bacterial evolution. Microbiol Rev 51: 221–271.
|