|
A lo largo de su
extensa costa las olas han recortado acantilados y estas paredes de
roca muestran sus estratos diseccionados como en un diorama de
museo. Acostumbraba a pasar mis vacaciones infantiles en un lugar
llamado Chapman's Pool en la costa del condado de Dorset; allí los
acantilados negros y sombríos de los esquistos de Kimmeridge se
encuentran moteados por ammonites y otros fósiles, blancos como la
nieve.
Por supuesto existen rocas más antiguas que
contienen microfósiles de bacterias, como las encontradas por Barghoorn y
Tyler, pero antes de que existieran los modernos métodos
de datación no se estaba seguro de su edad. El período con rocas más
antiguas que aquellas que contienen los fósiles visibles se llamó
Precámbrico porque era más distante en el tiempo que las rocas
cámbricas.
Escribiré este capítulo no como historia sino como un relato de la fisiología de un animal desconocido que vivió hace miles de años y acerca del cual sólo se dispone de una evidencia no mayor que unos pedazos de hueso, exactamente datados a partir de su contenido en isótopos de carbono. Mi interés principal se centra en los procesos geofisiológicos a gran escala que mantenían la Tierra en condiciones constantes y apta para la vida, y en el conocimiento de cómo operaban.
Con los huesos de la Tierra en la mente, consideraré un elemento importante, el calcio, y su papel crucial en todas las cosas vivas, desde nosotros a Gaia. Continuaré dedicando mi atención a aquellos otros elementos importantes, oxígeno, carbono e hidrógeno, con su capacidad de regulación y su efecto sobre él clima.
Este capítulo también tratará sobre la geofisiología de los
océanos, en particular el difícil problema de si la salinidad total
es mantenida sólo por fuerzas químicas y físicas o si es una
«maquinación» por parte de Gaia. Aunque el marco de éste capítulo es
el Proterozoico, la edad media de la Tierra, la mayoría de aspectos
discutidos no son exclusivos de este período, también actuaron en el
Arcaico y continúan actuando en el período presente.
Así como las fronteras políticas a menudo fallan en circunscribir regiones étnicas de manera precisa, los límites únicamente basados en consideraciones geológicas no siempre se adaptan al interés de la geofisiología. Como geofisiólogo prefiero poner mis marcas en el tiempo en que el ambiente se hizo predominantemente oxidante -o para expresarlo más profesionalmente-, en la transición de un ambiente dominado por moléculas dadoras de electrones, como el metano, a uno dominado por aceptadoras de electrones, como el oxígeno.
Como suele ocurrir, la
incertidumbre acerca de los acontecimientos de hace 2,5 eones es
grande y en adelante asumiremos que los registros geológicos y geofisiológicos definen el mismo período de la historia de la
Tierra.
La aparición de estas características sexuales secundarias es la respuesta a un flujo creciente de hormona pituitaria. Este acontecimiento principal puede localizarse en un tiempo definido, pero las características secundarias se extienden de forma arbitraria en el tiempo.
Entre el
Arcaico y el Proterozoico, la aparición del oxígeno como gas
dominante en la atmósfera fue el acontecimiento principal que marcó
un cambio profundo en el estado geofisiologico de la Tierra. El
resultado y las manifestaciones secundarias de este cambio -la
aparición de una nueva superficie, de una nueva química de la
atmósfera y de nuevos ecosistemas- es probable que se haya
distribuido durante un intervalo amplio de tiempo, y que ocurriera
en diferentes lugares y en diferentes momentos.
El Arcaico, cuyo ambiente estaba
lleno de moléculas dadoras de electrones (esto es, agentes
reductores), acabó encapsulado, como una región separada que existe
sólo cuando el oxígeno no está presente. La sumisión de los sistemas anóxicos al dominio de los oxidantes fue algo así como la conquista
por los normandos, de los primitivos sajones, que fueron relegados a
una posición subordinada -las clases bajas- de la cual, se dice a
menudo, no han escapado nunca.
Probablemente había un poco de oxígeno presente incluso a principios del Arcaico, del mismo modo que hay metano en nuestra atmósfera actual. La diferencia entre el aire arcaico y el aire proterozoico no era una simple cuestión de presencia o ausencia de oxígeno, sino de tendencias netas. En el Proterozoico una bicicleta desechada en aguas someras se hubiera oxidado dando óxido férrico insoluble que se depositaría en el fondo del mar. En el Arcaico se hubiera diluido lentamente en forma de iones ferrosos, solubles en agua, y no hubiera dejado rastro.
Durante el tiempo en que el flujo de metano
sobrepasaba al de oxígeno, la atmósfera inferior sólo contendría
cantidades traza de oxígeno. Los océanos y las rocas de la
superficie, ricas en iones ferrosos y sulfuros que atrapaban el
oxígeno, habrían absorbido hasta entonces una proporción del aporte
de oxígeno de las cianobacterias de manera que el aire habría
quedado en un estado netamente anóxico durante la mayor parte del
período arcaico.
Ello puede haber ocurrido como resultado de una caída repentina del metano atmosférico. Este acontecimiento habría venido acompañado por un proceso de enfriamiento porque el metano y sus productos de descomposición son gases de efecto invernadero. También hay razones geofisiológicas que favorecen una transición definida de forma abrupta a un estado oxidante.
Una vez que el oxígeno fotosintético empezó a ser dominante en la atmósfera y los océanos, la acción de la luz solar sobre el oxígeno habría producido radicales hidroxilo que oxidan el metano del aire. También habría habido consumidores alimentándose de la materia orgánica antes de que ésta pudiera llegar a los sedimentos anóxicos, lo que habría privado a los metanógenos del material para la producción de sus emanaciones gaseosas. Esta es la receta para una realimentación positiva en contra del metano y en favor del oxígeno.
Es probable que estos
sucesos ocurrieran en forma brusca en lugar de gradual.
A la luz de esta
teoría, ¿podemos describir un planeta viviente en el Proterozoico?
¿Qué sistemas reguladores controlaban su funcionamiento?
Las conjeturas que a continuación se exponen sobre la regulación del clima, el oxígeno, la salinidad y otras propiedades del medio ambiente parten de este contexto geofisiológico, en otras palabras, se consideran como si la Tierra fuese un organismo vivo. No se pretende de ningún modo que esto constituya una teleología, pues no se supone que el biota lleva a cabo una predicción o una planificación en la regulación de la Tierra.
Lo que se considera es
cómo se puede desarrollar un sistema regulador global a partir de la
actividad local de los organismos. No es inverosímil imaginar que
alguna bacteria nueva evolucionase en su medio ambiente para
conformar un sistema capaz de cambiar la Tierra. Efectivamente, la
primera cianobacteria, progenitora del ecosistema que empleaba la
energía luminosa para producir materia orgánica y oxígeno, hizo
esto.
Es el tercer ión en abundancia en el agua de mar, después del sodio y el magnesio. Tendemos a pensar en el calcio como un elemento nutritivo porque es un componente estructural esencial de nuestros huesos y dientes. También es crucial en numerosos procesos fisiológicos internos desde la coagulación de la sangre a la división celular. Es esencial para la vida pero, paradójicamente, muy tóxico en su estado iónico.
Dentro de nuestras
células, una concentración de iones calcio que exceda unas pocas
partes por millón es letal, una toxicidad comparable a la del
cianuro; sin embargo, los iones de calcio se encuentran en el agua
de mar a niveles diez mil veces superiores.
Ello es particularmente cierto en el caso del calcio. Imaginemos una bacteria de los océanos primitivos capaz de convertir los iones de calcio, abundantes y solubles en el agua de su medio interno, en carbonato cálcico insoluble. Esta simple reacción habría reducido de forma efectiva la concentración dentro de la célula de los iones potencialmente tóxicos, aislando el calcio en una forma insoluble y segura. En caso de que el calcio se encontrara en exceso en los océanos, tal como usualmente ocurre, dicha capacidad hubiera aumentado las oportunidades de supervivencia de este organismo y de su progenie.
Estos organismos estarían en ventaja en comparación con organismos que simplemente tolerasen la presencia de calcio en exceso. En la zona fótica del océano abierto, el crecimiento de estos organismos habría dado lugar a la deposición de grandes masas de calcio en el fondo oceánico. La lluvia de «conchas marinas» microscópicas, llamadas frústulos por los biólogos marinos, desde la superficie iluminada a las profundidades actúa como una cinta transportadora.
El alimento viaja hacia los consumidores de las
zonas inferiores, el océano se limpia y queda transparente, y
elementos potencialmente tóxicos como el cadmio se eliminan de las
regiones superficiales.
No veo cómo se
puede distinguir entre la cristalización espontánea de carbonato
cálcico sobresaturado en el Arcaico y la nucleación inducida por los
organismos. Creo que la nucleación de estados supersaturados y otros
estados meta-estables en la Naturaleza es un proceso geofisiológico
clave y que empezó a producirse en el Arcaico.
El proceso de
precipitación del carbonato cálcico fue un invento incluso más
grande, no solamente conllevó la regulación del calcio, del dióxido
de carbono y del clima sino también la gran ingeniería de las
estructuras de carbonato cálcico (los estromatolitos). Más adelante,
estos mismos procesos evolucionaron de manera que nuestras células
disponen de mecanismos intrincados por los que el calcio se deposita
en los huesos y dientes.
Según él, en algún tiempo lejano en la historia de la Tierra, se depositó una cantidad de caliza suficiente como para alterar la composición química de las rocas de la corteza del fondo oceánico cerca de los márgenes continentales. Ello dio lugar al suceso denominado por los geólogos transición basalto-ecoglita. Esta transición habría alterado tanto las propiedades físicas de las rocas de la corteza que habría posibilitado que la gran maquinaria del movimiento de placas empezara a girar.
Don Anderson comentó en su artículo en Science en 1984:
Para mí esta idea es muy interesante, pero admito que la mayoría de
los geólogos la encuentra extremadamente improbable. El fenómeno
podría haber sido desencadenado por la actividad de algunos
organismos capaces de separar una solución diluida de bicarbonato
cálcico en creta y dióxido de carbono y así evitar el envenenamiento
por calcio.
¿Ha sido por casualidad que los océanos se han mantenido siempre por debajo de este límite? ¿O ha sido la evolución estrechamente acoplada de la vida y el medio ambiente lo que dio lugar a una regulación automática de la salinidad de los océanos?
A menudo se dice que el medio salino interno óptimo para los organismos vivos, que es sorprendentemente similar en una gran variedad de ellos, refleja la composición de los océanos cuando empezó la vida. Es cierto que la salinidad de la sangre de las ballenas, personas, ratones y la mayoría de peces, tanto si viven en el océano como en agua dulce, es la misma. Incluso el fluido circulante del Artemia, un pequeño crustáceo que vive en soluciones saturadas de sal, comparte con nosotros la misma salinidad interna.
Pero en mi opinión esto no representa ninguna evidencia de la
salinidad de los océanos arcaicos, de la misma manera que los
niveles de oxígeno que ahora respiran estos organismos no dan
ninguna indicación de la abundancia de oxígeno en los orígenes de la
vida.
Uno lo puede comprobar por sí mismo haciendo burbujas de jabón con soluciones salinas de concentración creciente. Por encima de un 10 por ciento de sal no se producen burbujas. Ello es debido a que las moléculas de jabón consisten en una larga cadena de átomos de carbono fuertemente enlazados y rodeada de átomos de hidrógeno enlazados de manera igualmente firme. Uno de los extremos termina en un átomo de carbono enlazado con dos átomos de oxígeno.
Cuando este extremo de la molécula se disuelve en agua ligeramente alcalina, como lo son todas las soluciones de jabón, adquiere una carga negativa. La carga eléctrica negativa hace que el grupo terminal atraiga moléculas de agua y arrastre la cadena de hidrocarburo graso e insoluble dentro de la solución. La sal consiste en iones cloruro negativos e iones sodio positivos. Cuando hay muchos de estos iones en solución, compiten con la carga negativa del jabón en la absorción de moléculas de agua.
En presencia de una cantidad suficiente de sal, el jabón se separa del agua como la nata.
Por ejemplo, las membranas de los glóbulos rojos de la sangre humana están compuestas de partes aproximadamente iguales de colesterol y lecitina (un fosfolipido), diversas proteínas y otras sustancias grasas. Los glóbulos rojos se mantendrán expuestos a disoluciones de sal de hasta 0,8 molar (4,7 por ciento en peso). Por encima de esta concentración la membrana se malogra, y a concentraciones por encima de 2,0 molar la célula se destruye en pocos segundos.
A mediados de
los cincuenta pude demostrar mediante experiencias directas que los
perjuicios empezaban cuando la lecitina de la membrana de los
glóbulos rojos se dispersaba en la disolución salina fuerte. Este
tipo de desperfectos por la sal parecen ser generales entre las
células vivas y se observan en células de los cinco reinos
existentes.
Tienen que gastar energía en bombear agua contra la diferencia de presión osmótica entre su interior y el mar. La presión es del orden de la necesaria para bombear agua a una altura de 140 m en contra de la gravedad, alrededor de unas mil veces más alta que la presión sanguínea de los seres humanos. Las ventajas de un medio interno de bajo contenido en sal deben de ser considerables para requerir semejante inversión. Para una célula pequeña o una bacteria, la regulación a escala individual es un lujo que está mucho más allá de sus posibilidades.
Y no sólo en lo concerniente a la salinidad. Consideremos la
temperatura, por ejemplo. Para que un microorganismo pudiera
mantener tan sólo un grado de diferencia respecto a sus alrededores
sería necesario el consumo de mucho más oxígeno y alimento que el
que puede atravesar su superficie. No sólo eso, sino que un
gradiente de un grado de temperatura a través de la membrana celular
generaría una presión osmótica termal de 56 atmósferas, mucho más de
lo que puede resistir una membrana celular.
Hay bacterias tolerantes a la sal, los halófilos, que viven precariamente en las regiones salinas de la Tierra. Estas bacterias han resuelto el problema directamente desarrollando evolutivamente una membrana de especial estructura que no se altera con la sal. Funciona, pero al precio de que estos organismos no pueden competir con la mayor parte de bacterias cuando la salinidad es normal. Se encuentran limitadas a sus hábitats remotos y raros, y dependen del resto de la biosfera para mantener la Tierra apta para ellos.
Son como aquellos excéntricos de nuestra
sociedad cuya supervivencia depende del sustento que les podemos
proporcionar pero que raramente podrían sobrevivir solos.
Alrededor
del 15 por ciento del peso seco de las algas costeras es betaína, lo
que representa una desviación considerable de energía que en caso
contrario estaría disponible para el organismo. Por tanto, es
evidente que mantener los océanos tan diluidos como sea posible
-ciertamente mantenerlos lejos de la concentración crítica de 0,8
molar- representa una ventaja para el biota.
El único modo de eliminar las
enormes masas de sal de los océanos sería el aislamiento de agua
oceánica en lagunas seguido de la evaporación posterior del agua.
Esto habría requerido la edificación de grandes arrecifes de caliza
para atrapar la sal en lagunas evaporíticas. La magnitud total de
estos arrecifes dejaría pequeña cualquier construcción humana
concebible. Incluso es posible que el proceso de formación de
lagunas haya sido favorecido por el plegamiento de las rocas en los
márgenes continentales como consecuencia del movimiento de las
placas tectónicas.
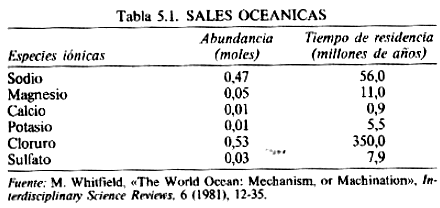
Estos se comportan como dos entidades se- paradas e independientes. Tanto el ión sodio como los otros iones positivos (potasio, magnesio y calcio) tienen tiempos de residencia relativamente cortos en el océano. Son eliminados por procesos químicos y bioquímicos, y también por reacciones químicas hidrotermales en las dorsales oceánicas.
Su destino final es la deposición en forma de sedimentos, arcillas, calizas y dolomitas. Básicamente, el problema de la salinidad para el biota es de hecho el problema de desembarazarse de los iones cloruro negativos.
Desde el punto de vista químico, el cloruro es bastante parecido a un átomo de argón, que es un gas totalmente inerte. Es una molécula esférica, escurridiza y lisa con escasa o nula tendencia a unirse a algo. No hay ningún intercambio bioquímico significativo con el cloruro.
Unos pocos sistemas poco comunes pueden generar cloruro de metilo a partir de sal, pero el ciclo de este compuesto es demasiado poco importante como para afectar a la salinidad del océano. Además, el cloruro presente en la molécula de cloruro de metilo pronto es reconvertido en ión cloruro y es arrastrado por la lluvia volviendo al mar. Los iones cloruro son eliminados de los océanos físicamente, mediante la transferencia de agua de mar a lagunas evaporíticas.
Sin ninguna planificación o previsión, los constituyentes de aquellas estructuras vivas, los estromatolitos calizos, se concentrarían en la costa y a la larga formarían lagunas en las que el agua de mar se evaporaría progresivamente depositándose sal. Al principio la formación de arrecifes sólo habría tenido un efecto local pero, a lo largo del tiempo, la mera presencia de la masa de caliza, habría empezado a afectar la corteza deformable de la superficie terrestre hundiéndola, y aumentando así el tamaño de las lagunas.
Siempre habría nuevos constructores de roca que
colonizarían la superficie de un arrecife a medida que el terreno se
hundiese, lo que tendería a mantener intacta la laguna.
Sin embargo, también hay nuevas lagunas evaporíticas formándose continuamente.
Los
cristales de sal en o cerca de la superficie están protegidos con su
propio barniz contra la disolución espontánea por el agua de lluvia.
Todos los pasos, desde la disminución individual de iones de calcio dentro de las células de un organismo vivo hasta los movimientos de las placas tectónicas, tienden a mejorar el medio ambiente para los organismos responsables de los mismos.
Sin embargo las relaciones entre biomineralización, acumulación salina y tectónica de placas son tan débiles que la mayoría de científicos piensan que se encuentran relacionadas aleatoriamente y no geofisiológicamente.
Tengo que continuar preguntándome sobre los límites de la manipulación gaiana e investigar el criterio correcto a partir de la pregunta
Quizás es improbable, pero recordemos que se ha dispuesto de miles de millones de años para el desarrollo de innovaciones geofisiológicas y su examen por selección natural. Deberíamos considerar la posibilidad de que este largo período hubiera sido suficiente para afinar la tosca geología en una geofisiología uniformemente regulada.
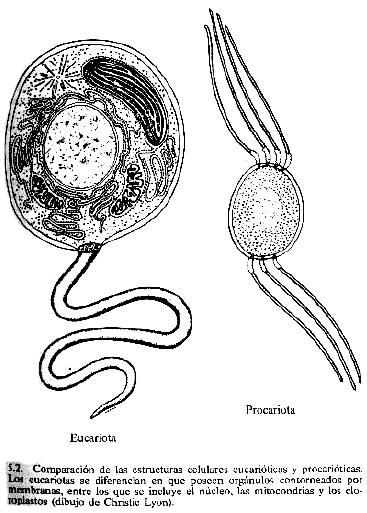
Pero en
poco tiempo aparecerían consumidores, organismos que aprendieron a
comer alimentos frescos y que ingerían a los fotosintetizadores. Las
células no tienen boca pero pueden ingerir otras células
englobándolas en una cavidad de su membrana, un proceso llamado
fagocitosis. La cavidad penetra en el interior de la célula y luego
se disuelve, dejando al cautivo atrapado dentro.
Aunque tratada superficialmente por
los biólogos del siglo XIX, el reconocimiento de esta asociación
poderosa de orgánulos trabajando dentro de las células en un proceso
de simbiosis, llamado endosimbiosis, se considera una contribución
de Lynn Margulis más que de ningún otro científico. Los
endosimbiontes ampliaron y expandieron las posibilidades de gestión
planetaria del biota y constituyeron la característica principal de
la historia de la Tierra durante el Proterozoico.
Una cuestión no resuelta acerca del proterozoico es cuál era la
concentración de oxígeno. ¿Permaneció alrededor del 0,1 al 1 por
ciento o aumentó hasta los niveles actuales o incluso más?
Ello es debido a que en una atmósfera de oxígeno sólo pueden existir trazas de hidrógeno o gases que lleven hidrógeno, como el metano. La única excepción es el agua, que no puede oxidarse más, y queda confinada a la atmósfera inferior por las bajas temperaturas que existen en la base de la estratosfera. Ahí queda literalmente congelada, y la parte superior de la atmósfera sólo contiene unas pocas partes por millón de vapor de agua.
La tasa actual de fuga de
hidrógeno al espacio se encuentra limitada por la sequedad del aire
superior y es de sólo 300.000 toneladas por año. Ello es equivalente
a casi 3 millones de toneladas de agua y dejaría detrás un exceso de
2,5 millones de toneladas de oxígeno. Parece mucho, pero una pérdida
de agua de esta magnitud habría eliminado menos del uno por ciento
de los océanos a lo largo de toda la existencia de la Tierra.
Una parte del carbono de los sedimentos interviene en la reducción de los sulfatos a sulfuros. La deposición de sulfuros también deja un incremento neto de oxígeno en el aire. El carbono y los sulfuros quedan incluidos en las arcillas y calizas sedimentarias. La sedimentación puede llevarse a cabo de manera que se formen combustibles fósiles, carbón y petróleo, aunque éstos sólo representan una pequeña proporción del carbono total y del azufre de los sedimentos. La deposición de todo este material oxidable es como un préstamo en la cuenta del oxígeno.
La deuda no se presenta mientras el carbono está enterrado o perdido
en el interior dé la Tierra, lo que permite que quede oxígeno libre
en circulación en el aire.
En parte, la elevada tasa actual de fotosíntesis se debe seguramente al reciclado rápido de carbono por los consumidores que respiran oxígeno. Estos metabolizan el 97,5 por ciento de los productos fotosintéticos, dejando sólo el 2,5 por ciento para los anaerobios.
En el Proterozoico había consumidores
que se alimentaban de materia orgánica y utilizaban el oxígeno para
metabolizarla. Su actividad probablemente haya sido menor que ahora,
pero mayor que en el Arcaico.
Entonces, ¿qué es lo que determinaba el nivel de oxígeno en el aire?
Desde el punto de vista de la geofisiología podemos suponer que la toxicidad inherente al oxígeno no fue totalmente vencida por los sistemas antioxidantes y por los enzimas de los organismos del sector aeróbico. En estas circunstancias el oxígeno mismo podría haber establecido su propio límite inferior y un límite superior para la mayor parte de los seres vivientes.
Tales propiedades pueden
estar geofisiológicamente reguladas.
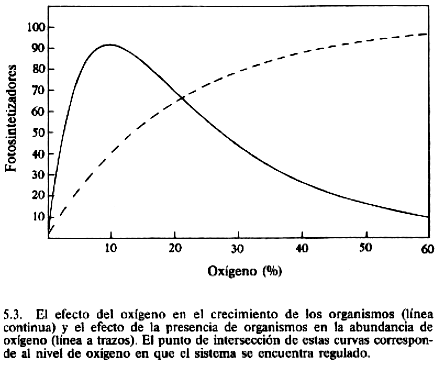
La línea de trazos indica la relación entre la población
del ecosistema aeróbico y el nivel estacionario de oxígeno; a más fotosintetizadores más oxígeno. Las dos curvas tienen un punto de
intersección en donde el nivel de oxígeno sería mantenido en
homeostasis por el sistema.
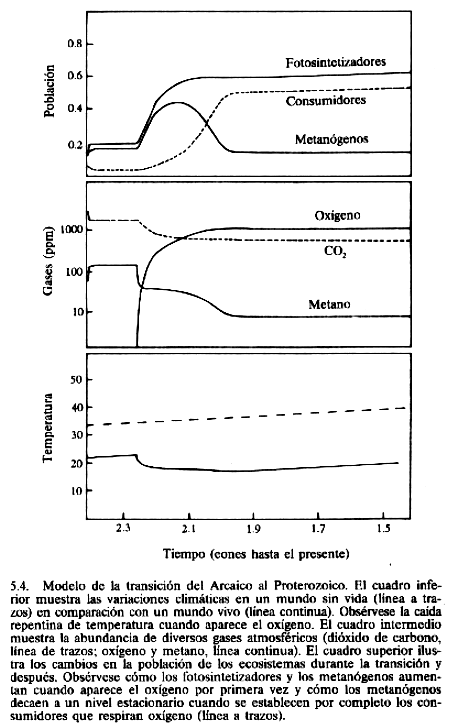
Entonces el oxígeno aumenta hasta que las propiedades geofisiológicas del sistema establecen un nuevo nivel estacionario en el que la abundancia de oxígeno se mantiene constante por un balance entre la cantidad de carbono enterrada y la cantidad de material reductor expuesto. El cuadro inferior ilustra las variaciones de temperatura planetaria en comparación con las de un planeta sin vida de la misma composición. El cuadro intermedio muestra la variación de los gases oxígeno, dióxido de carbono y metano.
El cuadro superior muestra los niveles de población de las
diferentes formas de vida.
El modelo de la figura 5.4 establece que, a largo plazo, la cantidad del carbono producido por los fotosintetizadores acumulada en los sedimentos es constante y que esta fuente de oxígeno también es constante. El proceso daría lugar a un agotamiento de la capacidad de absorción de oxígeno en las aguas y sedimentos anóxicos (serían oxidados), con lo que al final del Arcaico la concentración de oxígeno en la atmósfera subiría abundantemente.
La presencia de un exceso de oxígeno habría incrementado la tasa de meteorización y por tanto el aporte de nutrientes que a su vez habría posibilitado un ecosistema mayor. Ello redundaría en la sedimentación de más carbono y la subsiguiente subida de oxígeno. La presencia de este gas en la atmósfera se habría incrementado hasta que su toxicidad determinase un límite ambiental para los organismos vivos.
Esta situación limite produciría una reducción del sector anaerobio en el que se deposita el carbono, que se reduciría a un tamaño equivalente al del Arcaico, y la producción y pérdida de oxígeno se equilibrarían por la exposición de sustancias oxidables durante la meteorización.
En cierto sentido los ecosistemas aeróbicos existían ya en los albores de Gaia, desde el momento en que las primeras cianobacterias convirtieron luz solar en energía química de alto potencial y fueron capaces de sintetizar compuestos orgánicos y oxígeno a partir de agua y dióxido de carbono.
A medida que se extendían, estas cianobacterias habrían ocupado siempre una posición superficial para disfrutar y alimentarse de la luz solar.
Los sistemas anóxicos,
aquéllos cuyo alimento eran los cadáveres y los productos de las
cianobacterias, habrían proliferado de modo natural por debajo de
los fotosintetizadores para aprovecharse de la lluvia de alimento
desde arriba. Desde el principio habría existido una segregación
entre estos dos ecosistemas, y un gradiente de concentración de
oxígeno que disminuiría desde la región de su producción.
El bucle
de realimentación entre dióxido de carbono y clima estabilizaría el
sistema de forma adicional. Una vez superada la crisis del oxígeno
el Proterozoico pudo ser un tiempo confortable para Gaia, a
excepción hecha de la molestia persistente de los asteroides. El
nivel natural de dióxido de carbono habría proporcionado un clima
agradable y no se necesitaría un gran esfuerzo para regularlo.
El uranio natural siempre tiene la misma composición isotópica: 99,27 por ciento de uranio 238, 0,72 por ciento de uranio 235 y trazas de uranio 234. Sólo el uranio 235 puede actuar en las reacciones en cadena necesarias para la producción de energía o para las explosiones. Naturalmente este isótopo fisionable es vigilado celosamente y su proporción en el uranio se examina amplia y repetidamente. Imaginemos la sorpresa con que debieron encontrarse en la agencia francesa de energía atómica cuando se descubrió que la partida de uranio tenía una proporción mucho más pequeña de uranio 235 que lo normal.
¿Algún grupo clandestino en África o Francia había encontrado la manera de extraer el isótopo potencialmente fisionable y lo estaba utilizando para un uso terrorista con armas nucleares?
En un principio cualquier explicación siniestra parecía
probable. Cuando se encontró la verdad no sólo consistió en un
asunto científico fascinante sino que debió representar un alivio
inmenso para las mentes turbadas con imágenes de toneladas de uranio
235 puro en manos de fanáticos.
La concentración de las disoluciones de uranio no habría sido
superior a unas pocas partes por millón, y el uranio no habría sido
más que otro ión en solución. En lo que ahora es Oklo esta corriente
de agua fluía por un tapiz algal que incluía microorganismos con una
extraña capacidad para captar y concentrar uranio de forma
específica. Estos desarrollaron su tarea inconsciente tan bien que
al final se depositó suficiente óxido de uranio puro para iniciar
una reacción nuclear.
La reacción se estabiliza cuando los neutrones producidos compensan a los que escapan o son absorbidos por otros átomos. Este tipo de reactor no es explosivo, de hecho está autorregulado. Un aspecto fundamental del reactor es la presencia de agua, que tiene la función de retardar y reflejar neutrones. Cuando aumenta la producción dé energía el agua hierve y el reactor se frena. Una reacción de fisión nuclear es un tipo perverso de fuego, pues quema mejor cuando está bien empapado con agua.
Los reactores de Oklo funcionaron bien a la
escala de un kilovatio durante millones de años y utilizaron una
cantidad importante de uranio 235 para este fin.
La separación, casi
en estado puro, de sílice que realizan las diatomeas y de carbonato
cálcico que llevan a cabo los cocolitofóridos y otros organismos
vivos también constituyen ejemplos de este tipo de procesos que han
tenido un efecto profundo en la evolución de la Tierra.
En
la playa, el mar sería azul-grisáceo en lugar de marrón como en el
Arcaico. Tierra adentro, tras las dunas de arena y los guijarros,
estarían los tapices bacterianos, animados por algas verdes y
amarillo oro, que protegerían el sector anóxico donde se
encontrarían los lechos evaporíticos con sus capas profundas de sal
gema acumulada durante miles o incluso millones de años. Fuera, en
el mar, encontraríamos también las estructuras rocosas de los
estromatolitos.
La investigación
subsiguiente ha descubierto el hecho notorio de que los
microorganismos coralinos secretan una sustancia lipídica que forma
una capa monomolecular en la superficie del océano. Esta altera la
tensión superficial influyendo sobre las olas. Es interesante
especular sobre el desarrollo geofisiológico de esta interesante
propiedad de los microorganismos, preguntarse cuándo se desarrolló y
si es un mecanismo para protegerse de los embates del mar.
Estamos razonablemente seguros de que ninguno de ellos acabó con Gaia, de manera que una Gaia nueva hubiese tenido que renacer de las ruinas. La capacidad de recuperación tras perturbaciones grandes es una prueba de la salud de los sistemas geofisiológicos.
El hecho de que la vida haya persistido y se haya recuperado de tantas de estas catástrofes proporciona todavía más evidencia de la existencia de un sistema homeostático poderoso en la Tierra.
Los físicos y los geofisiólogos están completamente de acuerdo en que el dióxido de carbono atmosférico se encontraba a un nivel de alrededor del uno por ciento del volumen.
|