|
February 20, 1994 from EducateYourself Website
If you wish to look at living cells, the best research optical microscopes generally available throughout the world only reach about three thousand power. These microscopes in general cannot detect viruses, unless a fluorescence technique like Rife's fluorescence technique is used. These microscopes give very limited structural detail about living cell organelles. If the biologist wants detailed structural information about some cell structure, they use an electron microscope. However, the electron microscope picture is the picture of a dead, often highly degraded and distorted structure.
This is because the
sample preparation process, which produces a sample that can
withstand the conditions of high vacuum and bombardment by a high
energy electron beam has degraded and distorted the original living
structure. So at best you end up with a distorted snap shot of a non
living structure.
However, all this not withstanding. I also know the electron microscopes' limitations, both physically and in its actual use by researchers. If you have a interest in understanding biological microstructure, go to the trouble of going to a good research library and look up the Feb. 1944 issue of The Journal of the Franklin Institute or the 1944 Annual Report of the Board of Directors of the Smithsonian Institution.
In the RE. Seidel
and M. Elizabeth Winter article,
The New Microscopes, look at
the photographic plates. Note the high quality resolution comparable
to that of current electron microscope photographs. The photograph
of the typhoid bacillus was taken with the Rife Universal Microscope
at 23,000 power and then photographically enlarged to 300,000 power.
Further note that the resolution in this print is not as good as the resolution on the negative it came from do to the limitations in printing pictures in 1944 and even today. As was explained in technical detail in Appendix A, Rife had discovered an optical assembly configuration that effectively suppressed all Fraunhofer diffraction phenomenon. while at the same time he made the organism light itself by a natural fluorescence phenomenon.
This fluorescence phenomenon was achieved by illuminating the specimen with an intense narrow wavelength band of light. The particular band of light was unique to each microbe. Also note that this is a photograph of an intact living bacterium. If you are familiar with current microbiology. you know that little if any time is spent by the great majority of researchers watching and studying live microbes.
Except for spot optical
microscope checks to make sure live cultures are as they should be
or are as assumed they should be. research is carried out by
biochemical techniques the results of which are interpreted in the
light of past perceived research results. In short actually very
little live observation on microbe life cycles are carried out by
researchers anywhere on the entire planet.
After two years of reviewing the research for and against pleomorphism, it is clear that the monomorphists were wrong. The monomorphists won the argument because they had political prestige and economic positions of leverage. The monomorphists used optical microscopes and lab techniques not adequate to determine the issue due to inadequate magnification power, lack of non-lethal staining methods, sheer ignorance, and sloppy to lazy research work.
If you go to the trouble of looking up the Feb. 1944 issue of the Franklin Journal, note that the Rife microscope photograph of the typhoid bacillus clearly shows the formation of a filter passing form (the original operational meaning of the word virus) of the typhoid bacillus, in the top end of the bacillus. Rife found that when this bacillus virus was released by the bacillus, it had a bacterium flagella and was motile. Now all of this is just plain crazy, if you are a currently trained microbiologist.
However, no currently trained microbiologist owns or uses a Rife type optical microscope which could easily view this and the similar BX cancer virus, which is also a motile virus (ovoid body with bacterium flagella). The ovoid body dimensions of the BX cancer virus are 750 angstroms long by 500 angstroms thick. It is propelled by a proton transport flagella the same as the parent bacterium.
This "virus" will easily fit inside the so called AIDS virus (HIV) outer capsid and is comparable in size to the inner (HIV) capsid. I now ask you microbiologists reading this: Will this BX cancer "virus" be recognized in a high power electron microscope photograph for what it is or will it just be considered another piece of degraded cellular debris in the prepared cancer cell section sample? Much of what you see is what you are trained to see.
How are microbiologists
trained to see?
A one half centimeter on a side cube of carcinoma breast cancer tissue was placed into a test tube containing Kendall medium and incubated at 37 degrees centigrade. The test tube was then exposed to 24 hours of argon gas discharge light. The test tube growth medium was then examined under the Rife Universal microscope. at a magnification of 10.000 diameters. The medium was found to be teeming with animated ovoid microbes 1/15 microns long and 1/20 microns thick, which Rife eventually named the BX cancer virus.
This BX cancer virus was then carried through fourteen transplants from Kendall Medium to Kendall Medium. The animated BX cancer virus multiplied and remained of constant form. The fact that the BX cancer virus could multiply on a sterile non-living growth medium indicated that Rife's BX cancer "virus" was a living microorganism unlike the currently accepted understanding of a virus as a biological structure dependent on cellular metabolism to regenerate (multiply) and propagate its existence.
From current knowledge,
we must assume that Rife's BX cancer virus contains within its
structure, a gnome, DNA decoding enzymes, protein digestive enzymes,
transfer RNA, ribosomes, and associated proteins.
Rife found more than six forms, which the BX cancer virus could be transformed into.
These included:
Rife was not the only researcher to find a microbial cause for cancer. Many others have also. Nor was Rife the only one to build an optical microscope that could see the BX cancer virus.
Currently in Canada the biologist Gaston Naessens uses an ultraviolet microscope which can easily view the BX cancer virus in living blood from cancer patients. Naessens' microscope uses an ultraviolet light source which is first polarized. then focused down and sent through a frequency doubler crystal and finally sent into a special condenser section for dark field microscopy.
Looking at live blood
from cancer patients, Naessens has found and made videos of at least
sixteen different forms the BX cancer virus can be transformed into.
I have viewed some of these videos and the anti mated (motile)
BX and BY cancer virus are clearly visible and
look just as Rife described them.
On Sept. 10, 1953 The Washington Post reported the discoveries of Dr. Wheeler and her team from Rutgers-Presbyterian Hospital Laboratory which were disclosed at the 6th International Congress of Microbiology in Rome. They had found conducive proof of a microbial cause for cancer. When Dr. Wheeler and her group returned from Rome to Rutgers-Presbyterian Hospital they found that the funds for their laboratory were being cut off. The laboratory was closed.
This was the behind-the-scenes work and doings of Dr. Corneluis P. Rhoads, the head of Memorial Sloan-Kettering Cancer Center.
The fear of the cancer industry elite is and was immense. If the truth about the true cause of cancer becomes known, a cheap cure will be found shortly thereafter. This will kill the cancer goose which lays tens of billions of dollars worth of eggs a year.
Is there nothing these
scum will not do for their god money? No!
I will now share with you some observations about cancer cells and a classic experiment in which they are compared to normal cells, which suggests a simple answer to how cells infected with the BX cancer virus become cancerous.
It has long been noted that cancer cells act and appear somewhat like undifferentiated embryonic cells. Furthermore, cancer cells apparently have mostly an anaerobic (without oxygen) metabolism.
Note that the only time in the normal life cycle of mammalian cells in which they are of a undifferentiated embryonic nature and also have an apparent appreciable anaerobic metabolism is the period between the time the female egg, the ovum, has been fertilized in the fallopian tube and just before a viable placenta has developed in the uterus.
Geneticists and embryologists have shown that the entire development of the fetus from just-fertilized ovum to the fully developed fetus is governed completely by sequentially read and expressed genetic information.
There is an exceedingly complex genetic interchange and feedback control system in operation. Some of this genetic code is used only for a short period of time and is then sealed away not to be read or opened up again in the individuals existence, except during chromosome copying prior to cell division. Cancer cells act as though they have had some set of embryonic gene sequences reactivated.
However, in the now
mature differentiated mammalian cells from which this cancer cell
has been derived, the control system that normally would have
deactivated this embryonic gene sequence(s) is itself long since
deactivated. The cancer cell is in a run away catch 22 situation.
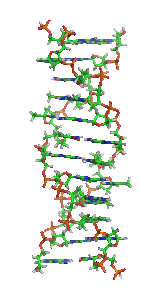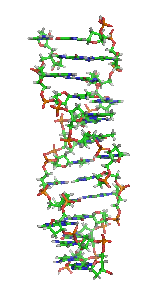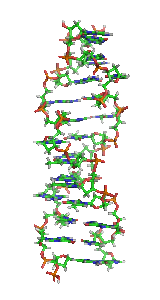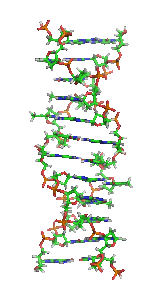
For this promoter protein to attach to its DNA coupling sequence at the beginning of the gene sequence, this sequence must be in the normal B-DNA right handed double helix form (see Figures 1 and 3).
If the coupling site code sequence or the DNA code sequence immediately in front of it has a blocking protein attached or is in the form of the left handed Z-DNA double helix (see Figure 2), the promoter protein can not bind/couple with its DNA code sequence and therefore the entire sequence of genes will not be read and expressed.
The Z-DNA double helix form is a very compact form of the double helix.
B-DNA double helix Z-DNA double helix
It has no major grove structure like the B-DNA double helix which allows a promoter protein to physically match up with a specific DNA code sequence which will manifest itself in the unique molecular structure of the surface of the major grove for that unique DNA code sequence.
The Z-DNA double helix structure gives very little information about what the DNA code sequence is in its core.
For a left handed Z-DNA double helix associated with a specific DNA code sequence to convert itself into a right handed B-DNA double helix, so that the promoter protein can attach, the concentrations of various ions in the cell nucleus must be in certain specific ranges for that specific Z-DNA sequence. The specific concentrations and ratios of ions in the nucleus is determined by the actions of ion gates and pumps in the cell outer membrane.
These ion gates and
pumps are controlled by messenger proteins and compounds from
both inside and outside the cell membrane. What this means is that
the cell genetic expression can be greatly influenced and controlled
by the genetic expression of other cells and cell sets (organs). And
of coarse during embryonic development this external cell influence
is in dominant control of the whole cell system of membrane ion
gates and pumps.
I believe the answer to all these questions is yes.
Of course there are many
other possibilities i.e. some of these protein fragments may act as
promoter proteins or combine with and remove blocker proteins,
thereby allowing a promoter protein to attach to a DNA sequence and
thereby initiate DNA transcription.
In 1948 Dr. Meryl S. Rose performed a mile stone experiment on salamanders. Rose transplanted frog kidney cancer tumor tissue onto a salamander's hind limb. These frog tumors were virus induced. The results of his experiment, however are the same even if the tumor is carcinogen induced, which was done later.
The transplanted tumors would grow and spread, leading to the salamander's death, if no intervention was taken. However, if Rose amputated the limb below or through the middle of the tumor, the salamander would regrow the limb and in the process the tumor(s) would disappear, even if the tumor had already spread to other body locations.
Tissue biopsies of the
wound region during regeneration showed that both salamander cells
as well as cancerous frog kidney cells dedifferentiated into
embryonic cell forms during the
blastema formation process as the
wound healed.
In later years researchers such as Becker demonstrated that it was the near unique ability of the salamander's nervous system to drastically change the ionic environment around blastema cells, along with hormone secretions from nerve dendrites, which allowed blastema cells to dedifferentiate into embryonic cells and then to red differentiate into the new cell types of the regenerating limb.
Becker and other researchers were able to get rats to regrow most of, or all of a amputated limb. They implanted a negative current source that produced a negative electric potential distribution inside the limb directly behind the amputation site. This closely mimicked what a salamander would have at that site if it were scaled up to the rats size.
To understand what is
happening here, you need to know that in a rat just as in a
salamander the myelin sheath cells coating the motor nerve fibers
carry an electron current through collagen fibers which are N-type
semi-conductors.
As a amputation wound heals over with skin, surface sensor nerve fibers cover over what is normally a motor nerve fiber region. In a short period of time the cells under the new forming skin layer can be converted into dedifferentiated embryonic cells under the influence or control of the external cell membrane ionic environment at the wound site as determined by the electric current-potential of the combined sensor and motor nerve sheaths activity in the wound area (blastema formation zone).
I can not here go into all of the wonderful detail of Becker's book. However, I hope I have given the reader at least an understanding of how cancer can possibly come about by a simple change in the ion environment in the cell nucleus. If you are interested in tissue regeneration or are a serious biologist. I can not recommend Becker's book enough. Particularly the last chapter, Postcript: Political Science.
This chapter with great
clarity and skill, clearly shows why we as a nation need to
dismantle all centralized cesspools of corruption as exemplified by
the National Institutes Of Health. The NIH needs to be
replaced by regional institutes which are government funded, but ran
and controlled by democratically elected administrators elected by
the research community.
This virus obtained in this manner was 100% effective in inducing cancer in lab animals. His form of the BX cancer virus was exceedingly virulent. Other researchers who apparently isolated the same BX cancer virus, or a form of it, and inoculated test animals by similar methods only had approximately 25% cancer induction rates.
A possible simple answer
for the discrepancy is that the ultraviolet light from the argon
discharge caused some of the adjacent thimine DNA base codes to
dimerize (chemically bond together). When the DNA reader enzyme
which translates the DNA base code into messenger RNA for protein
synthesis comes across a dimerized thimine base code pair, it stops
RNA synthesis. The reader enzyme then breaks into two fragments.
If this messenger RNA fragment contains the genetic RNA base code sequence for ribosome attachment, it will be read by the ribosomes and a protein fragment will be generated and released. In particular, if the RNA fragment is fed into a cluster of ribosomes (polyribosomes) which are located on or associated with the intercellular matrix web intersections, we can expect many copies of the coded protein fragment to be generated and released.
Furthermore, since the RNA fragment does not contain the normal stop synthesis code and message RNA end sequence base code, the RNA fragment is not likely to be immediately dismantled after polyribosome reading and protein synthesis like regular messenger RNA is. This fragment is likely to be read over again and again.
Now, if the generated
protein fragment happens to be an activator or suppressor of a cell
membrane ion channel or ion pump you have the potential beginnings
of a cancer producing situation as discussed above.
All of this is not the normal "plan" of the normal cell metabolism.
An excellent example of this sort of defective protein production and its cancerous consequences is the genetic disease xeroderma pigmentosum. In it the individual has an inherited defect in their ability to repair the aforementioned DNA base code dimerization damage.
They are hypersensitive to sun light exposure and develop pre-cancerous and cancerous skin conditions. They usually die of skin cancer before their twentieth birthday. Now what does this have to do with massive cellular tissue damage suffered by cancer patients while under going standard allopathic medical X-ray treatment for cancer?
As stated in Appendix B,
Rife's normal treatment for cancer patients was three minutes of
exposure once every three days to his frequency instrument.
The BX viruses disintegrated. In the normal carcinoma cancer cell, there are thousands of BX cancer viruses. When these BX cancer viruses all disintegrate together at the same time, they release their gnome, digestive enzymes, ribosomes, assorted proteinslenzymes, etc. into the cell. The cancer cell is overwhelmed, dies, and promptly disintegrates.
When using Rife's cancer
treatment method on a cancer patient that has undergone extensive
allopathic medical X-ray damage, there is the high possibility of an
encounter with a new kind of cancer cell which Rife's treatment
method won't work on.
All of this culminates in the possibility of a heavily radiation damaged BX cancer virus penetrating the cell membrane of a non cancerous cell and instigating production of cancer causing protein fragments as discussed above. But its own gnome so badly damaged that it can not propagate itself. If this were to occur, then a cancer cell could be created which was not infested with the BX cancer virus and therefore not treatable by Rife's frequency instrument or ultra sound of 11.789,000 or 23.560.000 cycles per second.
Of course the X-ray
radiation alone could generate a cancer cell that the original
Rife's treatment method would not cure.
Only by honest
researchers going back and looking at the suppressed results of past
honest cancer researchers can we hope to find honest valid answers
about cancer causes and cures.
P.S. - It is now empirically known that many types of cancer can be easily and quickly killed by exposure to pressure square waves of a frequency of approximately 2127 cycles per second.
It appears that one or more of the higher frequency hidden Fourier sine wave components. i.e.
...etc. , etc., opens
up ion gates on the cancer cells' membrane and radically changes the
ionic conditions inside the cancer cell such that it drops the
bi-lipid layer potential difference below some critical value below
which the cancer cell can not recover and it dies. |