|
La palabra teoría tiene la misma raíz griega que teatro. Ambas están relacionadas con la exhibición en un escenario.
Sin embargo, a los científicos muy trabajadores les gustaban las teorías repletas de predicciones que pueden ser comprobadas fácilmente. Tiene poca importancia si el punto de vista del teorizador es correcto o incorrecto: se estimula la investigación y la búsqueda, se descubren hechos nuevos, y se componen teorías nuevas.
Estar equivocado no supuso un demérito importante para la teoría de la creación continua de los astrónomos Hoyle, Bondi y Gold. Ahora la idea ha sido abandonada, pero en su día se trataba de un concepto intelectualmente satisfactorio. Las únicas teorías malas son aquellas que no pueden ser cuestionadas o evaluadas.
¿De qué sirve
una teoría que diga que el universo fue creado de forma completa,
con habitantes y todos con memoria de un pasado no existente, a las
15:37 horas el día 27 de octubre de 1917? No hay manera de probarlo
o rechazarlo, y no proporciona predicciones útiles.
Ellos nos han enseñado que se puede aprender mucho acerca de un
sistema a partir de pruebas no invasivas, que la vivisección no es
necesaria. Por tanto, la teoría de Gaia está abierta a varios modos
de investigación experimental.
Podemos seguir el
trayecto de los nutrientes esenciales desde las rocas al océano, al
aire y vuelta a empezar. También podemos apreciar cómo sistemas
diferentes pero interrelacionados se encuentran implicados en cada
paso.
Estos impactos liberan una cantidad de energía equivalente a
unos miles de veces la energía que sería liberada si todas las
cabezas nucleares estacionadas en los almacenes de armas actuales
explotasen al mismo tiempo. Estos acontecimientos producen algo más
que cráteres de 320 kilómetros; pueden destruir hasta el 90 por
ciento de todos los organismos vivos, desde los microscópicos hasta
los macroscópicos.
Si se duda de que la Tierra haya sido golpeada tan a menudo, basta con mirar el mapa de distribución de cráteres en las antiguas capas rocosas del Canadá (figura 3.1). Es como echar un vistazo a una región de la superficie lunar.
Sin embargo, en la mayor parte de las áreas continentales, y en toda la superficie del fondo del mar, la rapidez de los procesos de erosión allanadores y la expansión del fondo oceánico ocultan rápidamente la evidencia de todos los impactos, excepto los que corresponden a los más recientes.
No todos los acontecimientos catastróficos son producidos por causas externas; algunos, como la aparición de oxígeno en forma de gas, son generados por tensiones internas inherentes al sistema, y pueden ser comparados a las crisis de los organismos vivos, tales como la pubertad, la menopausia o la metamorfosis de una oruga en mariposa. El registro de las rocas, aunque difuminado por el tiempo y a menudo incompleto, todavía preserva alguna evidencia del estado físico y químico de la Tierra y de la distribución de especies antes y después de cada una de estas perturbaciones.
Sin embargo, desenredar el registro es como intentar encontrar trazas de la identidad de un terrorista a partir de los escombros del edificio que su bomba ha destruido.
Desde la certeza de sus conocimientos en biología dicen que nunca podría existir un organismo tan grande y, tal como lo ven, tan estable. Pienso que esta crítica es dogmática, y, como vimos en el capítulo anterior, fácilmente rebatible. El modelo simple del mundo de las margaritas ilustra cómo podría funcionar Gaia. Partíamos de un mundo imaginario que gira como la Tierra y que es calentado por una estrella gemela a nuestro Sol durante su movimiento orbital.
En este mundo, la competición por el territorio de dos especies de margaritas, una oscura y la otra clara, da lugar a una regulación exacta de la temperatura planetaria en el nivel apropiado para el desarrollo de estas plantas. No es necesaria ninguna previsión, ni planificación, ni propósito preestablecido. El mundo de las margaritas es una aproximación teórica a un planeta en homeostasis. Ahora podemos empezar a pensar en Gaia como una teoría, algo más que el simple «supongamos» de una hipótesis.
Cuando lo desarrollé comprobé que el número de especies que se puede incluir en él sólo parece estar limitado por la capacidad de los ordenadores utilizados y por la paciencia del investigador.
Independientemente de los detalles, los procesos de realimentación ambientales estabilizan el sistema de ecuaciones diferenciales que describe el crecimiento y la competición entre especies. Una gran parte de lo que sigue a continuación es el relato de mis exploraciones en el mundo de las margaritas, un informe de mis descubrimientos en él. He considerado que la mayoría de mis lectores no se sienten motivados por las expresiones matemáticas y, por tanto, no las incluyo. Para los que crean que cualquier teoría que no sea expresada en el lenguaje puro de las matemáticas es inadecuada, Andrew Watson y yo hemos descrito las matemáticas del mundo de estas plantas en nuestro trabajo en Tellus.
La estabilidad de dicho mundo no depende en ningún modo de
una selección precisa de condiciones iniciales, o de tasas de
crecimiento, y, tal como veremos en los últimos capítulos, el modelo
es general en su aplicación.
Tal como lo
expresaron, “las margaritas grises hubieran hecho trampa”.
Atribuí a las margaritas oscuras y claras una rebaja en la tasa de crecimiento de un uno (1%) por ciento por la fabricación de pigmentos. Estoy satisfecho de poder afirmar que el cínico parecer de los biólogos no se ve confirmado por este nuevo modelo. Así se puede observar en la figura 3.2. De nuevo se ha establecido que la luminosidad aumenta desde el 60 al 140 por ciento de la de nuestro Sol. Las poblaciones de margaritas se encuentran en el cuadro superior, con las oscuras a la izquierda, las grises en medio y las claras a la derecha.
El hecho de que las margaritas grises no gasten energía en la fabricación de pigmentos no les sirve de nada cuando su mundo es demasiado caliente o demasiado frío para que crezcan.
Pero las margaritas negras pueden florecer en el frío y las blancas
en el calor. Las margaritas grises proliferan cuando el clima es
templado y no se precisa regulación. En otras palabras, las
diferentes especies crecen cuando ellas y su medio ambiente están
adaptados el uno con el otro.
En la figura 3.3 se muestra la evolución de la temperatura, de las poblaciones de margaritas y otras propiedades de un mundo con veinte especies distintas. Como en el modelo de tres especies, se trata de un mundo cuya estrella se calienta a medida que envejece. El cuadro inferior muestra la evolución de la temperatura planetaria; el cuadro intermedio, la evolución de las poblaciones de las diferentes especies y el cuadro superior, la biomasa total y la diversidad del ecosistema representado en el modelo.
La diversidad del ecosistema es tanto mayor cuanto menor es la dificultad. Cuando el calor del Sol es exactamente el adecuado y no hace falta ningún esfuerzo para la regulación de la temperatura, entonces puede coexistir el mayor número de especies. Cuando el sistema se encuentra bajo alguna dificultad, cuando ha empezado a evolucionar o está agonizando, la diversidad es mínima y la población la constituyen casi enteramente las especies más oscuras o más claras.
Efectivamente, si alguna
especie está en situación ventajosa en esos períodos es la más
oscura o la más clara, pero nunca las grises.
El significado de las dos ecuaciones de este modelo puede expresarse como sigue: los zorros aumentan cuando el número de conejos crece, pero los conejos disminuyen cuando el de zorros aumenta. En esta relación hay un punto estable donde las dos especies coexisten en una proporción constante, excepto en el caso de una mala época que mate algunos conejos. Efectivamente, cualquier cambio de población que no provenga del interior del mismo modelo condena a este mundo simple a una fluctuación cíclica y nunca puede volver a una relación estable.
Observemos cómo ocurre.
Si una epidemia produce la muerte repentina de conejos, a continuación se producirá la muerte de zorros por falta de alimento. Los conejos se reproducen más rápidamente y pronto su población será superior al que había antes de la plaga. Pero ahora los zorros también empiezan a crecer, y la velocidad de crecimiento de los conejos se amortigua y disminuye cuando el incremento del número de zorros los amenaza.
Enseguida los conejos son demasiado escasos para alimentar a los
zorros, los zorros mueren y el ciclo empieza de nuevo.
Los ecólogos de campo han mostrado que en los ecosistemas simples se producen ciclos de poblaciones, pero al examinarlas más de cerca, nos parece que las observaciones de campo siempre se han realizado en ecosistemas enfermos o preparados por el hombre donde sólo unas pocas especies interaccionan entre sí, y donde sólo se consideran dos especies (por ejemplo plagas atacando la vegetación de un monocultivo agrícola, o enfermedades bacterianas de plantas y animales).
Estos ejemplos de dos especies, en las que las poblaciones siguen ciclos periódicos o fluctúan de una manera caótica e impredecible, pueden ser reproducidos con éxito por los sucesores matemáticos del famoso modelo de zorros y conejos de Lotka y Volterra.
Lo que estos modelos ecológicos y la propia teoría ecológica como ciencia han sido incapaces de explicar es la gran estabilidad de los ecosistemas naturales complejos como las selvas tropicales o, en otras palabras, resolver el acertijo de Darwin:
Los ecólogos han intentado vencer los inconvenientes de sus modelos sencillos incluyendo jerarquías estructuradas de especies que llaman «redes tróficas». En estas jerarquías hay una pirámide en cuya cúspide se encuentra el predador más alto, como por ejemplo el León, que presenta el menor número de individuos. Las poblaciones aumentan a medida que se desciende en cada nivel «trófico», hasta la base de la pirámide en que se encuentran los productores primarios más numerosos, las plantas, que proporcionan alimento para todo el sistema.
A pesar de los años de esfuerzo y el tiempo empleado en el cálculo, los ecólogos no han hecho ningún progreso real en el modelo teórico de un ecosistema natural complejo, como una selva húmeda tropical o el ecosistema tridimensional del océano.
Ningún modelo diseñado desde la ecología teórica puede explicar en términos matemáticos la estabilidad manifiesta de sistemas naturales tan amplios.
En efecto, el distinguido ecólogo, Robert May, en su libro Theoretical Ecology [Ecología teórica], escribe en el capítulo titulado «Modelos de ecosistemas con varias especies»:
May sigue escribiendo:
Esta negación reconoce la estabilidad de los ecosistemas complejos en el mundo real. Sin embargo, deja la impresión de que la diversidad es, en general, una desventaja, y que la naturaleza, no teniendo en cuenta las elegantes matemáticas de la biología teórica, de alguna manera ha hecho trampa.
Entonces, ¿qué es lo que confiere la gran estabilidad y ausencia de cualquier comportamiento cíclico o caótico a los modelos del mundo de las margaritas?
La respuesta es que las margaritas nunca pueden crecer de manera incontrolada; si lo hacen, el ambiente se hace desfavorable y se acota el crecimiento. De manera semejante, el medio ambiente no puede desplazarse hacia estados desfavorables en presencia de las margaritas; el crecimiento en respuesta de las margaritas del color apropiado lo evita. Es el acoplamiento estrecho entre estas relaciones, que condiciona tanto el crecimiento de las margaritas como la temperatura planetaria, lo que hace que el modelo funcione.
Quizá nuestra experiencia puede representarse mediante la
metáfora de que la familia y la sociedad funcionan mejor cuando
existen reglas firmes, pero aplicadas en su justa medida, que cuando
existe una libertad ilimitada.
Sorprendentemente, ni la adición de herbívoros ni las plagas afectan seriamente a la capacidad de las margaritas para regular el clima. Durante el curso normal de la evolución todas las poblaciones son estables y se recuperan pronto de las perturbaciones de las epidemias. Finalmente, el sistema ya no puede compensar el siempre creciente flujo de energía solar y se pierde. Como es de esperar, cuanto más cerca se encuentra del final más grande es el efecto de las perturbaciones.
Por ejemplo, las selvas de los trópicos húmedos están generalmente bien provistas de agua y sombra gracias a su cubierta de nubes; durante su existencia nunca se encuentran sometidas a una sequía prolongada como en una región desértica.
Por el contrario, los ecólogos teóricos ignoran el medio ambiente físico y químico. Para ellos el medio ambiente significa el conjunto mismo de especies, y un nicho es una parte de territorio negociado entre las especies como si uno pudiera mirar indistintamente el medio ambiente de Suiza para comprender la gente de Italia, Francia y Alemania. Desde este punto de vista las perturbaciones consisten en competencia o guerras.
Los trópicos húmedos han permanecido arbolados
a pesar de cambios en el clima de la Tierra que en términos humanos
pueden ser considerados como grandes, pero que son triviales a
escala planetaria. La presencia de una gran diversidad de especies
ayuda a esta gran capacidad para soportar el cambio climático.
Así, en la figura 3.5 se muestra un mundo de margaritas con 10 especies coloreadas cuya existencia estable es alterada de pronto por una epidemia que mata el 60 por ciento de las margaritas sin reparar en el color.
En el cuadro inferior, la línea a trazos representa la temperatura planetaria en ausencia de margaritas, a 40 °C se encuentra el límite para la vida. La línea continua muestra el clima con las margaritas presentes antes, durante y después de la perturbación. La temperatura se mantiene a unos 25 °C excepto cuando la epidemia golpea por primera vez la población de margaritas.
Cuando la perturbación cede el sistema rápidamente recupera el statu
quo. El cuadro superior muestra la variación en la distribución de
especies antes, durante y después de la catástrofe.
La respuesta rápida del mundo de las margaritas al cambio requiere una realimentación positiva e implica el crecimiento explosivo de las especies cuya interacción con el clima es más favorable.
Este modelo contiene 10 especies de margaritas distribuidos en un intervalo de colores uniforme y fijo desde el oscuro al claro.
Las mutaciones y la posibilidad de cambios evolutivos entre las especies constituyen un factor adicional a tener en cuenta. En este sentido, el cambio abrupto de la distribución de especies observado en el momento de la perturbación y de nuevo al final de la misma refleja la intensidad de la presión de selección en estos momentos. El modelo está fundamentalmente de acuerdo con las observaciones de Stephen Jay Gould y Niles Eldredge sobre la evolución puntuada.
En lugar de cambios estables y graduales, como en el punto de vista darwiniano convencional, hay períodos de evolución abrupta y rápida: las puntuaciones. La teoría de Gaia esperaría que la evolución del medio ambiente químico y físico y de las especies se produjera conjuntamente.
Habría períodos largos de homeostasis, con cambios pequeños en el medio ambiente y en la distribución de las especies, que serían interrumpidos por cambios súbitos de ambos órdenes.
Estos períodos puntuales podrían ser originados internamente por la evolución de algunas especies poderosas, como el hombre, cuya presencia altera el medio ambiente, o el resultado de influencias externas tales como el impacto de asteroides.
Este
experimento ilustra de qué modo la estabilidad, medida como la
capacidad para regular el clima, está correlacionada con la
diversidad. Las fluctuaciones de temperatura son máximas en los
momentos cercanos al nacimiento y la muerte del mundo de las
margaritas, cuando el número de especies es mínimo. En la flor de la
vida, el efecto de las perturbaciones se resiste de forma casi
completa.
Las perturbaciones
relativamente pequeñas de la oscilación orbital de la Tierra
producen pequeñas variaciones en la cantidad de calor recibida por
el Sol que resultan amplificadas debido a la inestabilidad de un
sistema moribundo. Estos argumentos no son necesariamente aplicables
a las glaciaciones ocurridas en un pasado más remoto, probablemente
debidas a causas diferentes.
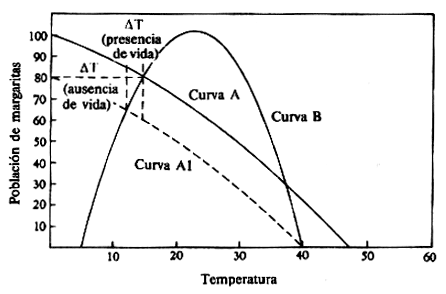
Este modelo está basado en un mundo de margaritas que sólo contienen margaritas blancas. Como son más claras de color que el suelo en el que viven tienden a incrementar el albedo de su localidad que, en consecuencia, es más fría que las áreas de superficie desnuda. Las margaritas influirán en la temperatura media del planeta desde las zonas que recubran. La relación entre el área cubierta por margaritas blancas y la temperatura superficial media del planeta se muestra mediante la curva A.
La línea de trazos paralela (A1)
muestra cómo se puede desplazar la relación si hubiese algún cambio
en alguna variable externa que influyese en la temperatura
planetaria -por ejemplo si la estrella que calentaba el mundo de las
margaritas disminuyese su aporte de energía radiante.
Las curvas A y B relacionan la temperatura planetaria con la población del mundo de las margaritas en un estado estacionario, y el estado estacionario de todo el sistema viene indicado por el punto de intersección de estas dos curvas.
En el ejemplo puede
observarse que hay dos soluciones estacionarias posibles. Resulta
que las soluciones estables se encuentran en las intersecciones
donde las tasas de cambio de la población y la temperatura tienen
direcciones opuestas. En términos matemáticos, cuando las derivadas
de las dos curvas tienen signos opuestos. La otra intersección no da
lugar a una solución estable. Si el mundo de las margaritas empieza
en alguna temperatura arbitraria, pero tolerable, se desplazará
hacia el punto de intersección superior y se asentará allí.
Supongamos, por ejemplo, que
la estrella se calienta tal como nuestro Sol lo hace. Si la
población de margaritas se mantuviese constante de manera
artificial, la temperatura planetaria acusaría simplemente el cambio
en el flujo de calor de la estrella; sobrevendría un cambio de
temperatura mucho más importante que si se deja crecer las
margaritas hasta su nuevo estado estacionario natural, desde donde
se opondrían al efecto provocado por el cambio en la radiación.
La hipótesis de que el crecimiento
de las margaritas está limitado a un margen estrecho de temperaturas
es crucial para el funcionamiento del mecanismo, pero esta
limitación dentro de un margen estrecho también se observa en toda
especie viva. La curva de crecimiento con punto óptimo (B) también
se observa en otras variables además de la temperatura, por ejemplo
el pH (puede ser muy ácido o muy alcalino, pero es preferible la
neutralidad). La mayoría de nutrientes también experimenta
restricciones similares; demasiado es veneno, demasiado poco produce
muerte por inanición.
Todos ellos mantenían el límite de 5 a 40 °C, pero comprendían una variedad de tipos que iban desde uno rectangular (un crecimiento constante a todas las temperaturas) a uno triangular (un incremento lineal hasta un máximo y luego una pérdida lineal de la misma pendiente). También se escogieron relaciones semicirculares y rómbicas. La única limitación resultante de estos cálculos consistió en la inestabilidad de los modelos cuando aparecía una parte horizontal en las relaciones.
Volvamos atrás a la figura 3.7 e imaginemos que la curva parabólica se sustituye por una en forma de sombrero recto. La sección horizontal, o parte superior del sombrero, implicaría la existencia de un crecimiento constante a cualquier temperatura dentro del intervalo de 5 a 40 °C, en consecuencia no podría haber ninguna regulación de la temperatura. Estos modelos son parecidos a los modelos de competencia simple de la biología de poblaciones que, al ignorarse el medio ambiente, resultan notoriamente inestables.
En la medida que la velocidad de
crecimiento de las margaritas varíe con la temperatura hasta un
máximo y luego decrezca, se obtiene un modelo que funciona, incluso
en los casos de curvatura suave en la parte superior del
semicírculo.
En su lenguaje, el mundo de las margaritas es un sistema de bucle cerrado. Los aparatos que no están autorregulados son a menudo inestables. Los ingenieros se refieren a ellos como de «bucle abierto», los bucles son los enlaces de realimentación entre las partes del sistema. Sin embargo, el mundo de las margaritas no tiene una forma idéntica a un aparato de ingeniería. Una diferencia clave consiste en la falta de «puntos fijos» (lo cual quizá también es característico de Gaia). En los sistemas manufacturados, el usuario fija la temperatura, la velocidad, la presión o cualquier otra variable.
El valor escogido es el punto fijo y el objetivo del
sistema es mantener ese valor si el medio ambiente externo cambia.
El mundo de las margaritas no tiene ningún objetivo establecido como
un punto fijo, sólo se sienta, como un gato, en una posición
confortable y resiste los intentos de desalojarlo.
En consecuencia sus modelos son incompletos, independientemente del grado de detalle que contengan. Es como si, en la figura 3.7, se considerase la biología de la relación entre población de margaritas y temperatura, pero sin referencia a la biofísica complementaria de las relaciones entre temperatura y población de margaritas. Un ingeniero o un fisiólogo identificarían que semejante planteamiento es un «bucle abierto», y en consecuencia de poco valor, excepto para estudiar casos extremos y patológicos.
También hay algo patológico en la arrogancia de algunos científicos que presumen de su conocimiento especial y de su desdén por otras disciplinas científicas. Hace sesenta años que el biólogo teórico americano Alfred Lotka, juicioso y de mente abierta, describió el modelo de competencia entre conejos y zorros en The Elements of Physical Biology [Los elementos de la biología física].
Desde entonces este modelo ha sido la inspiración de incontables investigadores en biología de poblaciones. Sin embargo ninguno hizo caso de su advertencia en la página 16:
Desde entonces, durante tres generaciones, los ecólogos teóricos han reproducido mediante modelos la evolución de los ecosistemas ignorando el medio ambiente físico, y tres generaciones de biogeoquímicos han reconstruido modelos de los ciclos de los elementos sin incluir siquiera los organismos como una parte dinámica y sensible del sistema.
En tiempos de Alfred Lotka la
resolución de sistemas de ecuaciones diferenciales no lineales era
una tarea amedrentadora, incluso para modelos simples como el mundo
de las margaritas. En la actualidad, con la disponibilidad de los
ordenadores no hay necesidad de circunscribirse a los límites
estrechos de una única disciplina científica.
Cuando están limitadas a una sola disciplina las matemáticas de los fenómenos naturales pueden convertirse en algo tan intrincado y complejo que a cada nuevo nivel investigado se abre un mundo nuevo de abstracción, todos ellos llenos de color. Es una pequeña maravilla que los usuarios de estas disciplinas imaginen que en sus mundos imaginarios vislumbran parte del mundo real mientras que, de hecho, están perdidos en el mundo fractal de un conjunto de Mandelbrot que reaparece siempre a cualquier nivel, desde menos a más infinito.
La ilusión es animada
por matemáticos profesionales que encuentran semejanzas entre sus
teorías matemáticas y las patologías del mundo real, y de numerosos
científicos matemáticos actuales cuya contemplación de los demonios
del hiperespacio - los «atractores extraños» del caos - es mucho más
seductora que el mundo aburrido y viejo de la naturaleza.
La primera prueba fue la expedición Viking a Marte. Aquella exploración confirmó la predicción, hecha a partir de análisis atmosféricos mediante astronomía infrarroja, de que Marte era un planeta sin vida.
Michael Whitfield, Andrew Watson y yo predijimos que la regulación a largo plazo del dióxido de carbono y del clima transcurre mediante el control biológico de la erosión de las rocas. Estas y otras pruebas se describen en los siguientes capítulos. Tiene poca importancia si la teoría de Gaia es correcta o equivocada; ya está proporcionando un punto de vista nuevo y más productivo acerca de la Tierra y de los demás planetas.
La teoría de Gaia conlleva una visión de la Tierra en la que:
Este último punto supone, además, una razón poderosa que justifica nuestra cólera instintiva sobre la desconsiderada supresión de especies, una respuesta a los que dicen que es algo puramente sentimental. Ya no tenemos que justificar la preservación de la rica variedad de especies en los ecosistemas naturales, como los de las selvas tropicales húmedas, mediante débiles razones humanistas como que, por ejemplo, podrían contener plantas con principios activos capaces de curar enfermedades humanas.
La teoría de Gaia nos hace cuestionar si estos ecosistemas ofrecen mucho más que esto. Mediante su capacidad de evaporar grandes volúmenes de vapor de agua a través de la superficie de sus hojas, los árboles sirven para mantener húmedos los ecosistemas de los trópicos y el planeta frío, proporcionando una sombra de nubes blancas reflectantes.
Su sustitución por áreas de cultivo podría precipitar un desastre regional de consecuencias globales.
|